Кроветворение
Содержание:
Постэмбриональный гемопоэз
Постэмбриональный гемопоэз представляет собой процесс физиологической регенерации крови, который компенсирует физиологическое разрушение дифференцированных клеток. Он подразделяется на миелопоэз и лимфопоэз.
Миелопоэз происходит в миелоидной ткани, расположенной в эпифизах трубчатых и полостях многих губчатых костей. Здесь развиваются эритроциты, гранулоциты, моноциты, тромбоциты, а также предшественники лимфоцитов. В миелоидной ткани находятся стволовые клетки крови и соединительной ткани. Предшественники лимфоцитов постепенно мигрируют и заселяют тимус, селезенку, лимфоузлы и некоторые другие органы.
Лимфопоэз происходит в лимфоидной ткани, которая имеет несколько разновидностей, представленных в тимусе, селезенке, лимфоузлах. Она выполняет функции образования T- и B-лимфоцитов и иммуноцитов (например, плазмоцитов).
Миелоидная и лимфоидная ткани являются разновидностями соединительной ткани, т.е. относятся к тканям внутренней среды. В них представлены две основные клеточные линии — клетки ретикулярной ткани и гемопоэтические клетки.
Ретикулярные, а также жировые, тучные и остеогенные клетки вместе с межклеточным веществом формируют микроокружение для гемопоэтических элементов. Структуры микроокружения и гемопоэтические клетки функционируют в неразрывной связи друг с другом. Микроокружение оказывает воздействие на дифференцировку клеток крови (при контакте с их рецепторами или путем выделения специфических факторов).
Таким образом, для миелоидной и всех разновидностей лимфоидной ткани характерно наличие стромальных и гемопоэтических элементов, образующих единое функциональное целое.
СКК относятся к самоподдерживающейся популяции клеток. Они редко делятся. Выявление СКК стало возможным при применении метода образования клеточных колоний – потомков одной стволовой клетки.
Пролиферативную активность СКК регулируют колониестимулирующие факторы (КСФ), различные виды интерлейкинов (ИЛ-3 и др.).
Каждая СКК в эксперименте или лабораторном исследовании образует одну колонию и называется колониеобразующей единицей (сокращенно КОЕ, CFU).
Исследование клеточного состава колоний позволило выявить две линии их дифференцировки.
Одна линия дает начало мультипотентной клетке — родоначальнице гранулоцитарного, эритроцитарного, моноцитарного и мегакариоцитарного рядов гемопоэза (сокращенно КОЕ-ГЭММ).
Вторая линия дает начало мультипотентной клетке — родоначальнице лимфопоэза (КОЕ-Л).
Из мультипотентных клеток дифференцируются олигопотентные (КОЕ-ГМ) и унипотентные родоначальные клетки.
Методом колониеобразования определены родоначальные унипотентные клетки для моноцитов (КОЕ-М), нейтрофильных гранулоцитов (КОЕ-Гн), эозинофилов (КОЕ-Эо), базофилов (КОЕ-Б), эритроцитов (БОЕ-Э и КОЕ-Э), мегакариоцитов (КОЕ-МГЦ), из которых образуются клетки-предшественники. В лимфопоэтическом ряду выделяют унипотентные клетки — предшественницы для B-лимфоцитов и для T-лимфоцитов. Полипотентные (плюрипотентные и мультипотентные), олигопотентные и унипотентные клетки морфологически не различаются.
Все приведенные выше стадии развития клеток составляют четыре основных класса, или компартмента, гемопоэза:
- I класс — СКК — стволовые клетки крови (плюрипотентные, полипотентные);
- II класс — КОЕ-ГЭММ и КОЕ-Л — коммитированные мультипотентные клетки (миелопоэза или лимфопоэза);
- III класс — КОЕ-М, КОЕ-Б и т.д. — коммитированные олигопотентные и унипотентные клетки;
- IV класс — клетки-предшественники (бласты, напр.: эритробласт, мегакариобласт и т.д.).
Сразу отметим, что оставшиеся два класса гемопоэза составляют созревающие клетки (V класс) и зрелые клетки крови (VI класс).
Эритропоэз у млекопитающих и человека протекает в костном мозге в особых морфофункциональных ассоциациях, получивших название эритробластических островков.
Эритробластический островок состоит из макрофага, окруженного одним или несколькими кольцами эритроидных клеток, развивающихся из унипотентной КОЕ-Э, вступившей в контакт с макрофагом. КОЕ-Э и образующиеся из нее клетки (от проэритробласта до ретикулоцита) удерживаются в контакте с макрофагом его рецепторами.
У взрослого организма потребность в эритроцитах обычно обеспечивается за счет усиленного размножения эритробластов. Но всякий раз, когда потребность организма в эритроцитах возрастает (например, при потере крови), эритробласты начинают развиваться из предшественников, а последние — из стволовых клеток.
В норме из костного мозга в кровь поступают только эритроциты и ретикулоциты.
Особенности дифференцировки гемопоэтических клеток
Кроветворная ткань костного мозга объединяет в своем составе комбинацию морфологически нераспознаваемых гемопоэтических клеток-предшественниц и клеток специфических рядов дифференцировки. Все кроветворные клетки, нераспознаваемые с точки зрения морфологии – это гемопоэтические стволовые клетки, которые могут быть:
- мультипотентными (дифференцируются во всех направлениях);
- полипотентными (развиваются только по некоторым из них);
- унипотентными (следуют только по определенному пути развития).
Другая часть клеток, которые удается распознать морфологически, формируется путем дифференцировки из более молодых предшественников, стремительно развивающихся дальше.
Миелопоэз может протекать в нескольких направлениях:
- мегакариоцитарное;
- эритроцитарное;
- моноцитарное;
- гранулоцитарное.
Лимфопоэз включает в себя две основные линии дифференцировки – образование лимфоцитов Т- и В-клеточного ряда. Каждая из них проходит в два этапа. Первый из них является антиген-независимым и заканчивается продукцией структурно зрелых, но иммунологически неактивных лимфоцитов. Следующий этап начинается после контакта с потенциальным антигеном и завершается выработкой специализированных иммунных клеток (Т-киллеров, Т-хелперов, Т-супрессоров, плазматических клеток, клеток памяти).
Каждый ряд дифференцировки кроветворных клеток дебютирует со стадии так называемых «бластов» (например, миелобласты). Для обозначения клеток промежуточного этапа используют приставку «про» и суффикс «цит» (например, проэритрокариоцит). Зрелые клеточные элементы имеют только суффикс «цит» (например, тромбоцит).
Следует отметить, что процесс дифференцировки различных видов клеточных элементов имеет свои особенности. Так, в гранулоцитарном ряде выделяют не одну, а несколько промежуточных стадий. В этом случае вслед за миелобластом образуется промиелоцит, затем миелоцит, метамиелоцит, а только после этого – зрелые клетки – эозинофилы, базофилы, нейтрофилы.
Кроме того, гемопоэз подразделяется на два периода:
- эмбриональный;
- постэмбриональный.
Эмбриональный период гемопоэза приводит к образованию крови как ткани и потому представляет собой гистогенез крови. Постэмбриональный гемопоэз представляет собой процесс физиологической регенерации крови как ткани.
Эмбриональный период гемопоэза осуществляется поэтапно, сменяя разные органы кроветворения. В соответствии с этим эмбриональный гемопоэз подразделяется на три этапа:
- желточный;
- гепато-тимусо-лиенальный;
- медулло-тимусо-лимфоидный.
Наиболее важными моментами желточного этапа являются:
- образование стволовых клеток крови;
- образование первичных кровеносных сосудов.
Несколько позже (на 3-ей неделе) начинают формироваться сосуды в мезенхиме тела зародыша, однако они являются пустыми щелевидными образованиями. Довольно скоро сосуды желточного мешка соединяются с сосудами тела зародыша, по этим сосудам стволовые клетки мигрируют в тело зародыша и заселяют закладки будущих кроветворных органов (в первую очередь печень), в которых затем и осуществляется кроветворение.
Гепато-тимусо—лиенальный
этап гемопоэза осуществляется в начале в печени, несколько позже в тимусе (вилочковой железе), а затем и в селезенке. В печени происходит (только экстраваскулярно) в основном миелоидное кроветворение, начиная с 5-ой недели и до конца 5-го месяца, а затем постепенно снижается и к концу эмбриогенеза полностью прекращается. Тимус закладывается на 7—8-й неделе, а несколько позже в нем начинается Т-лимфоцитопоэз, который продолжается до конца эмбриогенеза, а затем в постнатальном периоде до его инволюции (в 25—30 лет). Процесс образования Т-лимфоцитов в этот момент носит название антиген независимая дифференцировка. Селезенка закладывается на 4-й неделе, с 7—8 недели она заселяется стволовыми клетками и в ней начинается универсальное кроветворение, то есть и миелоилимфопоэз. Особенно активно кроветворение в селезенке протекает с 5-го по 7-ой месяцы внутриутробного развития плода, а затем миелоидное кроветворение постепенно угнетается и к концу эмбриогенеза (у человека) оно полностью прекращается. Лимфоидное же кроветворение сохраняется в селезенке до конца эмбриогенеза, а затем и в постэмбриональном периоде.
Следовательно, кроветворение на втором этапе в названных органах осуществляется почти одновременно, только экстраваскулярно, но его интенсивность и качественный состав в разных органах различны.
Медулло-тимусо-лимфоидный этап кроветворения
Закладка красного костного мозга начинается со 2-го месяца, кроветворение в нем начинается с 4-го месяца, а с 6-го месяца он является основным органом миелоидного и частично лимфоидного кроветворения, то есть является универсальным кроветворным органом. В то же время в тимусе, в селезенке и в лимфатических узлах осуществляется лимфоидное кроветворение. Если красный костный мозг не в состоянии удовлетворить возросшую потребность в форменных элементах крови (при кровотечении), то гемопоэтическая активность печени, селезенки может активизироваться — экстрамедуллярное кроветворение.
Постэмбриональный период кроветворения — осуществляется в красном костном мозге и лимфоидных органах (тимусе, селезенке, лимфатических узлах, миндалинах, лимфоидных фолликулах).
Сущность процесса кроветворения заключается в пролиферации и поэтапной дифференцировке стволовых клеток в зрелые форменные элементы крови.
СХЕМА КРОВЕТВОРЕНИЯ
Основные положения схемы кроветворения (отсутствие бессмертных «самоподдерживающихся» стволовых клеток, возможность сокращения числа митозов в процессе созревания, клональный характер кроветворения со сменой клонов, наличие ещё не выявленных, более ранних, чем стволовая клетка, предшественников и др.) были многократно подтверждены в течение многих лет. После утраты основного свойства стволовых и только стволовых клеток — способности к самоподдержанию — и невозможности определения самого понятия «стволовости», единственным критерием принадлежности клеток к стволовому отделу остается пролиферативный потенциал, достаточный для мультилинейного восстановления кроветворения после депрессии собственного кроветворения.
Время определения молекулярных основ биологии стволовых клеток ещё не пришло и составить более или менее ясную картину оркестровки генов в ходе кроветворных дифференцировок пока не удаётся.
В нижних этажах кроветворного дерева изменений очень немного. Помимо восьми ранее известных линий кроветворных дифференцировок выделены ещё новые: натуральные киллеры (клетки, участвующие в природном иммунитете, в том числе и против злокачественных клеток) и профессиональные антиген-презентирующие дендритные клетки. В настоящее время различают 11-12 линий дифференцировки кроветворных клеток.
Схему кроветворения (рис. 53-1) мы начинаем с единственного члена отдела тотипотентных предшественников, эмбриональной стволовой клетки (ЭС). Эта клетка способна к образованию клеток всех тканей организма. Эмбриональные стволовые клетки в нормальном развитии не существуют. Их выделяют из внутренней массы бластоциста на стадии примерно 100-120 клеток. В эмбриогенезе эти клетки быстро переходят на следующие стадии дифференцировки. Эмбриональные стволовые клетки образуются только в условиях остановки созревания клеток бластоциста вне организма, например при культивировании в определенных условиях. Дифференцировка клеток бластоциста блокируется и они способны пролиферировать практически бесконечно (больше 120 удвоений) без дифференцировки, без малигнизации, без изменений кариотипа и др. Снятие блока приводит к беспорядочной дифференцировке эмбриональной стволовой клетки. И хотя до разумного клинического применения эмбриональных стволовых клеток ещё далеко, уже сейчас целесообразно поместить этот предшественник в вершину кроветворной иерархии.
Васкуляризация. Иннервация. Возрастные изменения. Регенерация.
Васкуляризация. Костный мозг снабжается кровью посредством сосудов, проникающих через надкостницу в специальные отверстия в компактном веществе кости. Войдя в костный мозг, артерии разветвляются на восходящую и нисходящую ветви, от которых радиально отходят артериолы. Сначала они переходят в узкие капилляры (2—4 мкм), а затем в области эндоста продолжаются в широкие тонкостенные с щелевидными порами синусы (диаметром 10—14 мкм). Из синусов кровь собирается в центральную венулу. Постоянное зияние синусов и наличие щелей в эндотелиальном пласте обусловливаются тем, что в синусах гидростатическое давление несколько повышено, так как диаметр выносящей вены меньше по сравнению с диаметром артерии. К базальной мембране с наружной стороны прилежат адвентициальные клетки, которые, однако, не образуют сплошного слоя, что создает благоприятные условия для миграции клеток костного мозга в кровь. Меньшая часть крови проходит со стороны периоста в каналы остеонов, а затем в эндост и синус. По мере контакта с костной тканью кровь обогащается минеральными солями и регуляторами кроветворения.
Кровеносные сосуды составляют половину (50%) массы костного мозга, из них 30% приходится на синусы. В костном мозге разных костей человека артерии имеют толстую среднюю и адвентициальную оболочки, многочисленные тонкостенные вены, причем артерии и вены редко идут вместе, чаще врозь.
Капилляры бывают двух типов: узкие 6—20 мкм и широкие синусоидные (или синусы) диаметром 200—500 мкм. Узкие капилляры выполняют трофическую функцию, широкие являются местом дозревания эритроцитов и выхода в кровоток разных клеток крови. Капилляры выстланы эндотелиоцитами, лежащими на прерывистой базальной мембране.
Иннервация. В иннервации участвуют нервы сосудистых сплетений, нервы мышц и специальные нервные проводники к костному мозгу. Нервы проникают в костный мозг вместе с кровеносными сосудами через костные каналы. Далее покидают их и продолжаются как самостоятельные веточки в паренхиме в пределах ячеек губчатого вещества кости. Они ветвятся на тонкие волоконца, которые либо вновь вступают в контакт с костномозговыми сосудами и оканчиваются на их стенках, либо заканчиваются свободно среди клеток костного мозга.
Возрастные изменения. Красный костный мозг в детском возрасте заполняет эпифизы и диафизы трубчатых костей и находится в губчатом веществе плоских костей. Примерно в 12—18 лет красный костный мозг в диафизах замещается желтым. В старческом возрасте костный мозг (желтый и красный) приобретает слизистую консистенцию и тогда называется желатинозным костным мозгом. Следует отметить, что этот вид костного мозга может встречаться и в более раннем возрасте, например при развитии костей черепа и лица.
Регенерация. Красный костный мозг обладает высокой физиологической и репаративной регенерационной способностью. Источником образования гемопоэтических клеток являются стволовые клетки, находящиеся в тесном взаимодействии с ретикулярной стромальной тканью. Скорость регенерации костного мозга в значительной мере связана с микроокружением и специальными ростстимулирующими факторами гемопоэза.
Некоторые термины из практической медицины:
- миелограмма (миело- + греч. gramma запись) — выраженный в форме таблицы или диаграммы результат микроскопии мазка пунктата костного мозга, отражающий качественный и количественный состав ядросодержащих клеток миелоидной ткани.;
- миелоидная реакция (миело- + греч. —eides подобный) — появление в периферической крови малодифференцированных клеток, относящихся к грануло- и эритропоэтическому ряду; наблюдается при метастазах злокачественной опухоли в костный мозг, а также при сепсисе, туберкулезе и некоторых других болезнях;
- остеомиелит (osteomyelitis; остео- + греч. myelos костный мозг + ит; син.: костоеда — устар., паностит) — воспаление костного мозга, обычно распространяющееся на компактное и губчатое вещество кости и надкостницу;
| Часть первая – Общая характеристика, классификация; красный костный мозг | immunesys1.mp37 122 кБ |
| Часть вторая – Вилочковая железа — тимус | immunesys2.mp35 310 кБ |
| Часть третья – Селезенка | immunesys3.mp36 073 кБ |
| Часть четвертая – Лимфатические узлы, лимфоидная ткань слизистых оболочек | immunesys4.mp37 152 кБ |
Органы человека: костный мозг
Время созревания гранулоцитов в костном мозге составляет 60-200 часов, при этом в процессе дифференцировки морфологически распознаваемые клетки гранулоцитарного ряда проходят 4 митоза.
Родоначальницей всех зернистых лейкоцитов является миелобласт (клетка 4 класса).
Его размеры от 12 до 22 мкм. Миелобласты отличаются нежной структурой ядра, как правило, содержащего от 2 до 5 ядрышек. Цитоплазма различной степени базофилии, окружает ядро небольшим пояском. Цитоплазма содержит азурофильную (неспецифическую) зернистость, не всегда отчетливо видимую.
В результате митотического деления и одновременно дифференцировки миелобласты переходят в следующую стадию развития — промиелоциты (клетки 5 класса).
Его размеры 10-24 мкм.
Ядро занимает большую часть клетки, расположено эксцентрично. Форма ядра круглая или овальная. Цитоплазма базофильная, наряду с азурофильной грануляцией может появляться специальная — нейтрофильная, эозинофильная или базофильная.
Из промиелоцитов развиваются миелоциты (клетки 5 класса).
Миелоциты — клетки размером 10-18 мкм.
Ядро круглое или овальное, ядрышки отсутствуют. Цитоплазма содержит ту или иную специфическую зернистость — нейтрофильную, эозинофильную, базофильную. Ядерно-цитоплазматическое отношение сдвинуто в пользу ядра. Миелоциты среди гранулоцитов являются последними клетками, способными к делению. Дальнейшую дифференцировку гранулоциты проходят без деления в составе непролиферирующего пула костного мозга.
Следующей стадией созревания гранулоцитов являются метамиелоциты (клетки 5 класса)
.
Их размеры 10-15 мкм. Ядро имеет вид подковки или боба; структура ядра более грубая, чем у миелоцита. Цитоплазма нейтрофильного метамиелоцита окрашивается в розовый цвет, эозинофильного — бледно-голубой, базофильного — голубовато-фиолетовый.
В цитоплазме различают специфическую зернистость. Ядерно-цитоплазматическое отношение 1:1.
В костном мозге из метамиелоцитов образуются палочкоядерные лейкоциты (клетки 5 класса
).
Их размеры составляют 9-12 мкм. Ядро имеет вид палочки средней толщины (часто изогнутой в виде буквы S), грубой структуры. В цитоплазме различима специфическая зернистость. Ядерно-цитоплазматическое отношение сдвинуто уже в сторону цитоплазмы.
Последней стадией созревания являются сегментоядерные гранулоциты (клетки 6 класса
)
а)нейтрофилы
– имеют размеры 11-12 мкм.
Ядро состоит из нескольких сегментов (2-6). Цитоплазма содержит мелкую зернистость, окрашивающуюся нейтральными красками в фиолетовый цвет;
б) эозинофилы
имеют размер 12-13 мкм. Ядро эозинофила имеет чаще всего 2-3 крупных сегмента. Цитоплазма содержит крупную зернистость, окрашивающуюся эозином в розовый цвет;
в) базофилы
— имеют размер 9-10 мкм.
Ядро широкое, неправильной лопастовидной формы. Цитоплазма содержит крупную зернистость, окрашивающуюся основными красками в фиолетовый цвет, черно-синие тона. Различают 2 вида базофилов: циркулирующие в периферической крови — базофильные гранулоциты и локализованные в тканях — тучные клетки или тканевые базофилы.
Контроль гемоцитопоэза
Гемоцитопоэз объединяет процессы, связанные с преобразованием различных клеток в зрелые элементы крови. Так обеспечивается естественное сокращение лишнего количества форменных элементов. Полипотентные стволовые клетки самостоятельно проходят регенерацию. Они могут образовываться вне органов кроветворения. При стандартной дифференциации полипотентные клетки, сосредоточенные в костном мозге, способствуют зарождению зрелых элементов крови. Это:
- базофилы;
- эритроциты;
- различные типы лимфоцитов;
- эозинофилы;
- нейтрофилы.
К ним относят:
- попадание инфекции в организм;
- механические повреждения и другие факторы, способствующие потере определенного количества крови;
- адаптация к условиям высокогорья.
Как осуществляется кроветворение: механизмы
Процессы разрушения красных кровяных шариков и их образования строго сбалансированы. Если организм теряет какое-то количество крови, то не проходит 2—3 недель, как снова восстанавливается исходный уровень числа эритроцитов и концентрации гемоглобина. При этом всегда наблюдается значительное убыстрение образования красных кровяных телец (эритропоэза) в костном мозге.
Не вызывает сомнений факт существования в организме особых механизмов регуляции эритропоэза, хорошо выявляемых тогда, когда под влиянием каких-либо причин резко уменьшается количество эритроцитов и в связи с этим развивается кислородное голодание — гипоксия.
Законно предположить, что уменьшение снабжения организма кислородом автоматически приводит к увеличению продукции красных кровяных телец.
- Хорошо известно, что у жителей высокогорья, а так же у альпинистов, достигающих больших высот, число эритроцитов заметно повышается по сравнению с исходной нормой.
- И наоборот, если в барокамере создать повышенное давление кислорода, то через некоторое время можно отметить постепенное затухание, «вялость» красного кроветворения, вплоть до полного его прекращения.
Возникает вопрос о механизме «эритроцитостимулирующего» действия кислородного голодания. Большим количеством исследований установлено, что этот фактор убыстряет кроветворение через посредство особого вещества, стимулирующего эрицропоэз и получившего название «эритропоэтин».
В 1906 г. два французских исследователя — Карно и Дефляндер — обнаружили, что сыворотка крови, взятая у кроликов через 20 часов после массивной кровопотери и введенная другому здоровому кролику, способствовала у последнего приросту эритроцитов на 2—3 млн. в 1 мм3 крови, а также увеличению количества гемоглобина.
Последующие эксперименты показали, что кислородная недостаточность любого происхождения способна повышать эритростимулирующие свойства кровяной сыворотки.
Наиболее убедительные доказательства существования в организме стимулятора красного кроветворения были представлены в опытах на искусственно сращенных между собой (наподобие сиамских близнецов) крысах.
Этот интересный опыт выглядел так: одна из крыс дышала газовой смесью, содержащей пониженное количество кислорода, а ее партнер — воздухом с нормальным содержанием кислорода. И оказалось, что у обоих животных в костном мозге происходило одинаковое разрастание клеток «красного ряда», а в периферической крови — значительное увеличение эритроцитов.
Объяснить это можно следующим образом: у крысы под влиянием кислородного голодания образуется вещество эритростимулирующего действия, т. е. эритропоэтин, который переходит с кровью через сращенные кровеносные сосуды в организм партнера и вызывает у него активизацию кроветворения.
Что такое гемопоэз и последствия его нарушений
Кроветворение – это очень сложный процесс, он должен постоянно регулироваться и никак не изменяться. Любое изменение в ту или иную сторону приводит к нарушениям в организме и к возникновению различных заболеваний. Гемопоэз, что это такое и с чего он начинается, рассмотрим ниже.
В первые дни жизни эмбриона кроветворение происходит в стенках желчного мешка, в его утолщениях. Начало этого процесса приходится на 16-19 день развития, а после 60-го дня кроветворение начинает происходить в печени, селезенке и в тимусе. Затем, когда развился костный мозг (а он развивается самым последним из кроветворных органов), эти функции переходят к нему. Тогда в печени активное образование крови прекращается.
Как уже упоминалось выше, из стволовых клеток образуются миелоидные клетки (эритроциты, моноциты, тромбоциты, гранулоциты). Этот процесс получил название миелопоэз. Выработка предшественников лимфоидных клеток – лимфопоэз. Процесс миелопоэза осуществляется в миелоидной ткани, которая находится трубчатых, а также во многих губчатых костях. Во время миелопоэза происходит изменение типа клеток. Например, прежде чем стать тромбоцитами, меняется кариотип полиплоидных мегакариоцитов, а при образовании эритроцитов исчезает ядро у клеток эритробласт.
Селезенка, лимфатические узлы, тимус (вилочковая железа) и костный могз отвечают за протекание лимфопоэза. В лимфоидной ткани происходит образование лимфоцитов, плазмоцитов, а также удаляются клетки и остатки из распада.
При некоторых заболеваниях возникают нарушения в костном мозге, что приводит к нарушению кроветворения. Например, при увеличении количества тромбоцитов становится гуще кровь, что может привести к застою крови в различных органах, замедлению кровотока и образованию тромбов, что является большой опасностью для жизни человека.
Если же клеток крови вырабатывается недостаточно, то симптомы проявляются по мере того, как отмирают нормальные клетки. Самыми «маложивущими» являются гранулоциты, поэтому при их исчезновении организм становится более уязвимым для различных инфекций. Потом начинают убывать эритроциты, что проявляется в тромбоцитопении. Появляется кровоточивость. А при уменьшении количества эритроцитов характерными признаками являются бледность, общая слабость, одышка даже при, казалось бы, небольших нагрузках.
Познания в этой области улучшились только недавно, когда генная инженерия и культивирование клеток достигли нового уровня. Появилась возможность регулировать процесс кроветворения, подходя к этому индивидуально, в зависимости от того, сколько клеток недовырабатывается.
Следите за своим здоровьем! И будет не лишним узнать какие причины низкого гемоглобина в крови, и как бороться с проблемами такого характера.
Типы гематопоэза
Гематопоэз бывает нескольких типов, в зависимости от дифференцирующихся клеток:
- эритропоэз;
- гранулоцитопоэз;
- лимфоцитопоэз;
- моноцитопоэз;
- тромбоцитопоэз.
Эритропоэз
Процесс образования эритроцитов называется эритропоэз или эритроцитопоэз. Он начинается с дифференцировки плюрипотентной стволовой клетки в мультипотентную стволовую клетку, за которой следует колония эритроцитов, образующая клетку-предшественник и проэритробласт. Проэритробласт – это клетка, которая содержит ядро и множество рибосом, в которых начинает синтезироваться гемоглобин. Базофильный эритробласт, меньший по размеру, отличается от проэритробласта. После еще нескольких стадий деления образуется ретикулоцит, который является первой неядерной клеткой, содержащей остатки клеточных органелл. После того, как ретикулоцит теряет свои органеллы, образуется эритроцит. Зрелый эритроцит выполняет свои функции и живет около 120 дней. Затем он расщепляется на селезенку и гемоглобин, который содержался в нем, превращается в печень и выводится из организма в виде билирубина.
Гранулоцитопоэз
Гранулоцитопоэз начинается с плюрипотентной гемопоэтической стволовой клетки. Первым идентифицированным предшественником гранулоцитов является миелобласт. Миелобласт образует промиелоцит и миелоцит. Миелоциты содержат специфические гранулы и делятся на нейтрофилы, эозинофилы и базофилы. Нейтрофильный миелоцит имеет почечное ядро и два типа гранул (азурофильные и специфические). Эозинофильный миелоцит имеет овальное ядро и эозинофильные гранулы. Базофильный миелоцит имеет овальное ядро и азурофильные гранулы. После нескольких последовательных делений миелоцитов образуются метамиелоциты, которые снова делятся на нейтрофилы, эозинофилы и базофилы. Затем следует образование палочковидных и сегментарных гранулоцитов. Время, необходимое для дифференцировки стволовых клеток в зрелые гранулоциты, составляет около 10 дней.
Лимфоцитопоэз
Лимфоцитопоэз – это процесс образования лимфоцитов. Он проходит следующие стадии: гемопоэтические стволовые клетки, клетки-предшественники лимфоцитов, лимфобласты и лимфоциты. Три типа клеток развиваются из клеток-предшественников – B- и T-лимфоцитов и естественных клеток-киллеров (NK-клеток).
Моноцитопоэз
В результате моноцитопоэза образуются моноциты, которые попадают в кровоток и при необходимости превращаются в макрофаги. Процесс образования моноцитов начинается с гемопоэтических стволовых клеток, за которыми следует колония моноцитов, образующая клетки-предшественники, монобласты, промоноциты и моноциты. Время, необходимое для дифференциации зрелого моноцита, составляет около 55 часов.
Тромбоцитопоэз
В процессе тромбоцитопоэза образуются тромбоциты. Он начинается с гемопоэтических стволовых клеток, за которыми следуют колониеобразующие единицы мегакариоциты и тромбоциты. Мегакариоциты представляют собой крупные клетки, из которых протромбоциты образуются путем частичной фрагментации, из которой образуются тромбоциты.
Регуляция гемопоэза
Кроветворение регулируется:
- факторами роста, обеспечивающими пролиферацию и дифференцировку СКК и последующих стадий их развития,
- факторами транскрипции, влияющими на экспрессию генов, определяющих направление дифференцировки гемопоэтических клеток,
- витаминами, гормонами.
Факторы роста включают колониестимулирующие факторы (КСФ), интерлейкины и ингибирующие факторы. Они являются гликопротеинами, действующими и как циркулирующие гормоны, и как местные медиаторы, регулирующие гемопоэз и дифференцировку специфических типов клеток. Почти все факторы роста действуют на СКК, КОЕ, коммитированные и зрелые клетки. Однако отмечаются индивидуальные особенности действия этих факторов на клетки-мишени.
КСФ действуют на специфические клетки или группы клеток на различных стадиях дифференцировки. Например, фактор роста стволовых клеток влияет на пролиферацию и миграцию СКК в эмбриогенезе. В постнатальном периоде на гемопоэз оказывают влияние несколько КСФ, среди которых наиболее изучены факторы, стимулирующие развитие гранулоцитов и макрофагов (ГМ-КСФ, Г-КСФ, М-КСФ), а также интерлейкины.
Большинство указанных факторов выделено и применяется для лечения различных болезней. Для получения их используются биотехнологические методы.
Дифференцировка полипотентных клеток в унипотентные определяется действием ряда специфических факторов, поэтинов — эритропоэтинов (для эритробластов), гранулопоэтинов (для миелобластов), лимфопоэтинов (для лимфобластов), тромбопоэтинов (для мегакариобластов).
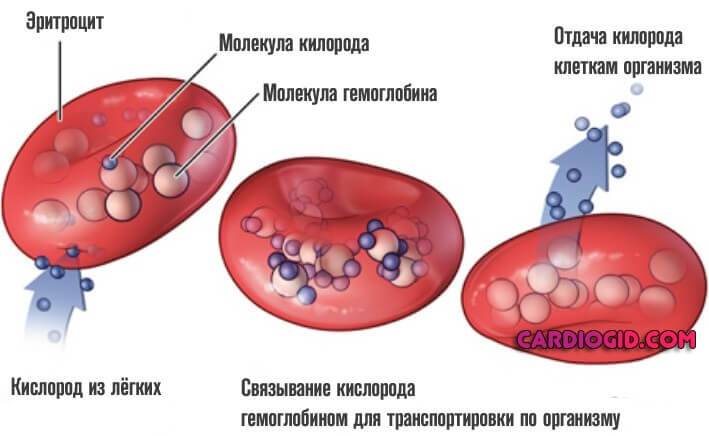
Большая часть эритропоэтина образуется в почках. Его образование регулируется содержанием в крови кислорода, которое зависит от количества циркулирующих в крови эритроцитов. Снижение числа эритроцитов и соответственно парциального давления кислорода, является сигналом для увеличения продукции эритропоэтина. Эритропоэтин действует на чувствительные к нему КОЕ-Э, стимулируя их пролиферацию и дифференцировку, что в конечном итоге приводит к повышению содержания в крови эритроцитов.
Тромбопоэтин синтезируется в печени, стимулирует пролиферацию КОЕ-МГЦ, их дифференцировку и образование тромбоцитов.
Ингибирующие факторы дают противоположный эффект, т.е. тормозят гемопоэз; их недостаток может быть одной из причин лейкемии, характеризующейся значительным увеличением числа лейкоцитов в крови. Выделен ингибирующий лейкемию фактор (ЛИФ), который тормозит пролиферацию и дифференцировку моноцитов-макрофагов.
Витамины необходимы для стимуляции пролиферации и дифференцировки гемопоэтических клеток. Витамин В12 поступает с пищей и соединяется с внутренним фактором (Касла), который синтезируется париетальными клетками желудка. Образуемый при этом комплекс, в присутствии ионов Са2+, соединяется с рецепторами эпителиоцитов подвздошной кишки и всасывается. При всасывании в эпителиоциты поступает лишь витамин В12, а внутренний фактор освобождается. Витамин В12 поступает с кровью в костный мозг, где влияет на гемопоэз, и в печень, где может депонироваться. Нарушение процесса всасывания при различных заболеваниях желудочно-кишечного тракта может служить причиной дефицита витамина В12 и нарушений в гемопоэзе.
Некоторые термины из практической медицины:
- анемия (син. малокровие) — состояние, характеризующееся снижением содержания гемоглобина в единице объема крови, чаще при одновременном уменьшении количества эритроцитов;
- анемия ахрестическая — общее название анемий, развивающихся вследствие неспособности эритробластов костного мозга использовать какие-либо антианемические факторы (цианокобаламин, фолиевую кислоту, железо и др.) при их нормальном поступлении в организм;
- хлороз ранний (син.: бледная немочь, хлороз ювенильный) — железодефицитная анемия у девушек в период полового созревания, проявляющаяся алебастровой бледностью кожи с зеленоватым оттенком, извращением вкуса, олигоменореей;
- хлороз поздний (син. анемия железодефицитная эссенциальная) — железодефицитная анемия у женщин в возрасте старше 30 лет, обычно связанная с маточными или другими кровотечениями, проявляющаяся извращением вкуса и обоняния, признаками атрофии слизистой оболочки полости рта и пищевода, выпадением волос, искривлением ногтей, позже — выраженными признаками анемии;
Что влияет на кроветворение?
Причины малокровия (анемии) многообразны. Самая распространенная из них — дефицит железа или витамина В12. Малокровие может сопутствовать многим заболеваниям. В связи с тем, что количество циркулирующей в теле крови установить довольно сложно (кроме того, дефицит жидкости организм восполняет путем ее изъятия из тканей), диагностика анемии основывается на оценке количественных показателей крови. Основным в анализе крови является содержание эритроцитов и лейкоцитов, а также концентрация гемоглобина.
Для стимуляции кроветворения (особенно выработки в красном костном мозге эритроцитов) врач назначает лекарства, в состав которых входят железо, кобальт, витамин В12, фолиевая кислота, а также печеночные экстракты.
Малокровие (анемия) — уменьшение в крови количества эритроцитов и гемоглобина. Основные симптомы: утомляемость, одышка, бледность кожи, плохая сопротивляемость инфекциям. Существует много причин возникновения анемии, например, значительная кровопотеря из-за травмы или в результате хронического кровотечения.
Дополнительно статьи на данную тему:
Строение и функции эритроцитов крови